Hi! Long time no see!
(I think we’ve reached the point where it’s weird to say happy new year. I could swear xkcd had a pertinent chart of funny, but I couldn’t find it.)
Once upon a time, I briefly mentioned the problematic relationships of hemichordates. Since a short paper bearing on the subject came out relatively recently (i.e. in December, yes, I’m far behind the times ;)), I thought I’d revisit it.
To begin, let’s orient ourselves on my trusty old animal phylogeny.
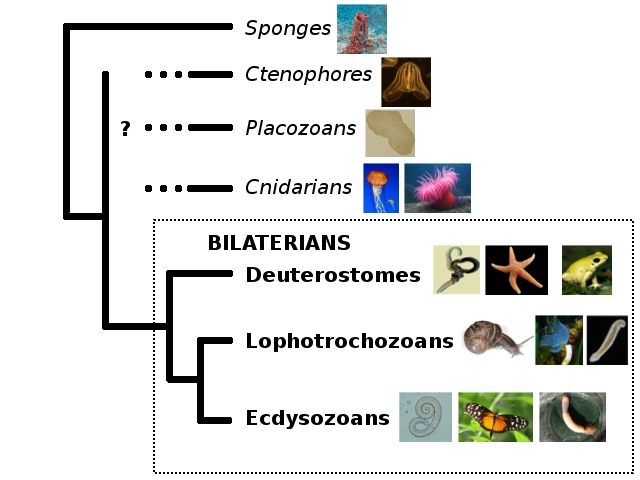
Hemichordates are a phylum of deuterostomes, and their closest relatives appear to be echinoderms like starfish. The inside of Deuterostomia looks something like this:

Hemichordates come in two flavours: the butt-ugly (but nevertheless intriguing) acorn worm, which even the artistic eye of 19th century zoologists couldn’t make appealing (a selection of them from Johann Wilhelm Spengel’s work below):

… and the slightly nicer-looking pterobranch. Well. They’re kind of fluffy. That counts as “nicer,” right? (A couple of Cephalodiscus from the Halanych lab below):

Acorn worms and pterobranchs have different bodies adapted to very different lifestyles. Pterobranchs are stalked, tentacled filter-feeders that often clone themselves into colonies that live together in a branching tube system. Acorn worms are solitary burrowers without tentacles, tubes or shells. Hemichordates possess features in common with vertebrates, such as gill slits, and they seem a lot less freakish than their sister phylum Echinodermata. So hemichordates are kind of the natural go-to group to look for properties of the deuterostome common ancestor.
The only problem is, to do that, you need a solid understanding of hemichordate phylogeny itself. Because there are two very different kinds of hemichordates, you have to first figure out which of those best represents their common ancestor: the sit-at-home plankton sifter or the roaming mud-eating worm. (Maybe neither. Wouldn’t that be funny.) And, as it happens, there’s some disagreement about that.
One view, espoused by the mighty zoological tome of Brusca and Brusca (2002) among others, puts acorn worms and pterobranchs as separate sister groups, and considers pterobranchs the more conservative of the two. The Bruscas write, on page 869, that “the enteropneusts [= acorn worms] have lost [their tentacles], no doubt in connection with their development of an infaunal lifestyle.” In this view, the deuterostome ancestor was a sessile filter feeder, and the long worm-like body and general moving-aboutiness of other deuterostomes is a new feature.
The other hypothesis, backed by DNA sequence data (Cannon et al., 2009)* and more recently the discovery of a tube-dwelling acorn worm from the Cambrian (Caron et al., 2013), is that pterobranchs are a weird subgroup of acorn worms and therefore unlikely to say much about our own distant ancestors.
One thing that AFAIK both camps agree on is that the ancestral acorn worm had a larva that looked nothing like an acorn worm. That’s something pretty common for marine invertebrates. Creatures as different as sea urchins and ragworms explore the seas by way of tiny, planktonic larvae that later metamorphose into a completely different animal**. (Tornaria larva of an unidentified hemichordate below by Alvaro E Migotto from the Cifonauta image database.)

However, the specific family of acorn worms that pterobranchs supposedly come from does not have such a larval stage. They develop more or less directly from fertilised eggs into mini-acorn worms.
Pterobranchs are poorly studied, so not much is known about their babies. Are they like the conventional acorn worm larva, with its distinctive body plan and curly rows of cilia? Or are they more straightforward precursors of the adult, like their presumed closest cousins? Stach (2013) describes a larva of the pterobranch Cephalodiscus gracilis that looks more like the latter. He found the minuscule creature crawling around in a colony of adult Cephalodiscus, and used thin sections and transmission electron microscopy to make a 3D reconstruction of it.
(His account of finding the baby makes me wonder how the hell he knew it did belong to Cephalodiscus. If my experience with tube-dwelling marine invertebrates is anything to go by, being found in a certain animal’s home is no guarantee that you’re related to said animal. I suppose, incomplete though they may be, older descriptions of pterobranch babies were good enough to identify the little guy?)
The image that emerges is of a rather featureless little sausage. According to Stach, it has a through gut, one full-fledged and one partially formed gill opening (asymmetry like that is not unheard of in deuterostome embryos/larvae), as well as a body cavity and a bunch of muscle cells. What it doesn’t have is any trace of the bands of cilia that “typical” acorn worm larvae use to swim and feed, nor some other structures (e.g. nerve centres) that characterise such larvae.
Taken at face value, this would suggest (assuming this is a typical pterobranch larva) that the pterobranchs-are-acorn worms people are right. I have my reservations, and not just because a sample size of one makes me statistically nervous. Using this description as evidence for evolutionary relationships assumes that traditional larvae with ciliary bands are hard to lose. But that’s quite possibly not the case.
Echinoderm larvae, for example, have changed a lot even in the last few million years. The changes occurred many times independently, and often involved a return from a full-fledged larval stage to more direct development (Raff and Byrne, 2006). I don’t know whether acorn worms display a similar sort of flexibility. How many have even been studied in terms of development?
So: detailed internal structure of a pterobranch larva? Cool. As to the worms first hypothesis… “consistent with” would be a better description than “supports”, I think.
***
Notes:
*Although microRNAs beg to differ (Peterson et al., 2013).
**The history of these larvae is a mighty can of worms, or trochophores and tornariae as the case may be. I shall say no more on the matter here. 🙂
***
References:
Brusca RC & Brusca GJ (2002) Invertebrates (second edition). Sinauer Associates.
Cannon JT et al. (2009) Molecular phylogeny of hemichordata, with updated status of deep-sea enteropneusts. Molecular Phylogenetics and Evolution 52:17-24
Caron J-B et al. (2013) Tubicolous enteropneusts from the Cambrian period. Nature 495:503-506
Peterson KJ et al. (2013) MicroRNAs support the monophyly of enteropneust hemichordates. Journal of Experimental Zoology B 320:368-374
Raff RA & Byrne M (2006) The active evolutionary lives of echinoderm larvae. Heredity 97:244-252
Stach T (2013) Larval anatomy of the pterobranch Cephalodiscus gracilis supports secondarily derived sessility concordant with molecular phylogenies. Naturwissenschaften 100:1187-1191
